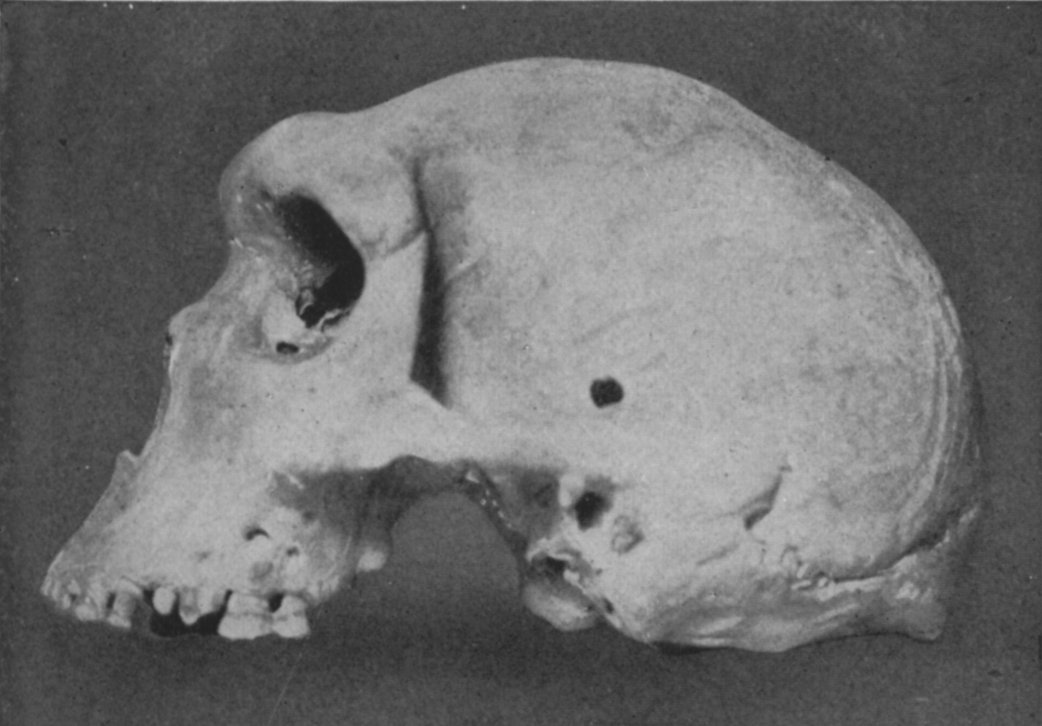
Skull
from Zambia, dated to 110,000 years ago. Modern humans co-existed with archaic
groups in Africa, particularly in the south and west.
When
and where did modern humans emerge? Anatomical evidence points to somewhere in
eastern Africa some 300,000 years ago. The time of origin is different if we
look at behavioral and genetic evidence. Sophisticated tool-making, detailed
artwork, and other signs of “behavioral modernity” appeared only 70,000 years
ago (Brown et al., 2012). Genetic evidence points to a series of demographic
expansions between 80,000 and 60,000 years ago in eastern Africa, with the last
one spreading throughout Africa and into Eurasia (Watson et al. 1997). At that
moment, an innovation in thinking seems to have given these truly modern humans
an edge over everyone else.
As
these humans spread throughout the world, to what extent did they intermix with
the more archaic groups they replaced? We can answer this question for Eurasia
by comparing the modern human genome with reconstructed genomes of the
now-extinct Neanderthals (Europe, Middle East, and Central Asia) and Denisovans
(East Asia, Southeast Asia). Present-day Eurasians have relatively low levels
of archaic admixture: about 2% from Neanderthals and up to 5% from Denisovans
(Sankararaman et al. 2016).
What
about Africa? Unfortunately, we have not yet reconstructed the genome of any
archaic population from that continent. We probably never will, given that DNA
tends to degrade quickly in tropical climates. In theory, there should be more
admixture in Africa than in Eurasia, since many archaic Africans would have
been "near-modern," i.e., much more similar in appearance, behavior,
and genetic makeup to modern humans than either Neanderthals or Denisovans.
Greater genetic similarity would have also made hybrid infertility less likely.
Indeed, it looks like male fertility suffered from hybridization with
Neanderthals or Denisovans, given that present-day humans have a lower
proportion of archaic ancestry on the X chromosome and in genes
disproportionately expressed in the testes (Sankararaman et al. 2016). In these
parts of the genome, natural selection has stepped in to remove archaic
admixture.
The
above speculations seem borne out by a recent and still unpublished paper. Its
authors, Sriram Sankararaman and Arun Durvasula, came up with a novel way to
measure admixture from an unknown archaic group, essentially by using a machine
learning algorithm (which they validated with data on Neanderthal introgression
in present-day Europeans). When they applied this method to Yoruba from
Nigeria, they found a level of archaic admixture higher than in any other human
population known to date:
Our results suggest that Yoruban individuals trace about 7.9% of their genomes to an as yet unidentified archaic population. This is in agreement with some results from previous papers in other African populations such as the Biaka and the Baka, suggesting that there was a rich diversity of hominin species within Africa and that introgression was commonplace. (Sankararaman and Durvasula 2018)
This finding is consistent with previous archaeological and genetic evidence, particularly from western and southern Africa. Both regions seem to have had archaic populations until recent times:
-
A skull from a Nigerian site (Iwo Eleru) is only about 16,300 years old and yet
looks intermediate in shape between modern humans on the one hand and
Neanderthals and Homo erectus on the
other. It resembles the skull of a near-modern human, like the Skhul-Qafzeh hominins
who lived in the Middle East some 80,000 to 100,000 years ago (Harvati et al.,
2011; Stojanowski, 2014).
-
Genomic analysis of 16 prehistoric Africans suggests that modern humans spread
out of eastern Africa and into western Africa, where they mixed with an archaic
population as divergent from modern humans as Neanderthals were, the time of
separation from modern humans being 200,000 to 300,000 years ago. This archaic
admixture is estimated at 9% in Yoruba and 13% in Mende (Skoglund et al. 2017)
-
Genomic analysis shows an apparently higher level of Neanderthal ancestry in
the Yoruba of Nigeria than in the Luhya of Kenya. This admixture seems to come from
a Neanderthal-like population that formerly lived in West Africa (Hawks 2012)
-
A skull from Zambia has been dated to 110,000 years ago and yet looks very much
like a Homo erectus (Bada et al.,
1974; Stringer, 2011).
- About 2% of the current African gene pool
comes from a population that split from ancestral modern humans some 700,000
years ago. This archaic DNA was then picked up by modern African humans about
35,000 years ago, probably in central Africa because this admixture is highest
in pygmy groups from that region (Hammer et al. 2011).
-
Genomic analysis of western African pygmies (Biaka and Baka) indicates
frequent, low-level interbreeding between archaic and modern humans, including
an admixture event within the last 30,000 years (Hsieh et al. 2016).
-
Jawbone fragments from South Africa exhibits significant size and morphological
variability, indicating admixture with an archaic population. The fragments
fall within the range of 110,000 to 60,000 years ago (Malekfar, 2012)
-
Sub-Saharan Africans exhibit dental traits that distinguish them from other
modern humans (Sub-Saharan African Dental Complex). These traits are shared
with extinct hominids and many extinct and extant nonhuman primates (Irish 1998).
When dentitions are compared from western, central, eastern, and southern Africans,
these ancestral traits appear to be least present in Kenyans and Tanzanians
(Irish 1998). The SSADC thus seems least present in the "homeland" of
modern humans (eastern Africa) and more present farther west and south.
Is the estimate of
7.9% archaic admixture a lower bound?
While
the new finding of 7.9% archaic admixture is higher than what we see in other
modern humans, the actual figure may be higher still. Sankararaman and
Durvasula attribute this 7.9% admixture to "a deeply-diverged archaic
population," while nonetheless acknowledging the "rich diversity of
hominin species within Africa." Dienekes (2018) likewise notes that
multiple admixture events had occurred between modern African humans and a
range of "Palaeoafrican" groups.
Thus,
Sankararaman and Durvasula are measuring admixture only from a highly divergent
archaic group, apparently the same one that Skoglund et al. (2017) found in
their study of the Yoruba. Indeed, the two studies found almost the same level
of archaic admixture in the Yoruba: 7.9% versus 9%. Although Sankararaman and
Durvasula validated their methodology with data on Neanderthal admixture in
Europe, the two situations are not really comparable. In Europe, modern humans
encountered only one archaic group over a relatively short time span,
intermixture taking place essentially between 60,000 and 50,000 years ago with
a second event more than 37,000 years ago (Yang and Fu 2018). In Africa, modern humans likely encountered a
range of archaic groups over a longer time, including "near-moderns"
whose ancestors diverged from those of modern humans less than 200,000 years
ago.
If
we include introgression from these “near-moderns,” the total for archaic admixture
in present-day sub-Saharan Africans should be much higher. Indeed, 13% of the sub-Saharan gene pool seems
to come from a demographic expansion that took place some 111,000 years ago and
which probably brought the Skhul-Qafzeh hominins to the Middle East (Watson et
al. 1997). Those hominins were anatomically modern, or almost so, but
culturally Neanderthal.
Did archaic
admixture help or hinder?
Mainstream
evolutionary theorists have argued that admixture does more harm than good. As Ernst
Mayr (1970, p. 80) wrote:
The claim has been made that species owe much of their genetic variability to introgressive hybridization. However, all the evidence contradicts this conclusion so far as animals is concerned. Not only are F1 hybrids between good species very rare, but where they occur the hybrids (even when not sterile) are demonstrably of inferior viability. The few genes that occasionally introgress into the parental species are not coadapted [...] and are selected against. Introgressive hybridization seems to be a negligible source of genetic variation in animals.
This view has been challenged by Hawks et al. (2007), who argue that gene introgression helped modern humans adapt to new environments. Instead of starting from scratch, they could cherry-pick genes that had already been tried and proven by the populations they were replacing:
Compared with novel mutations, archaic genetic variants would have had several qualities that, in some cases, may have enhanced their selective value. Because they had long existed within human populations, these alleles had a much lower chance of being strongly deleterious. [...] Alleles with local advantages might never have been selected within the expanding modern population until it reached new climatic regimens. The spread of modern humans may have attained a burst of evolutionary change by drawing on the fruits of the existing adaptations of archaic humans. (Hawks et al. 2007)
The latest findings seem to lie between the above two views. Introgression can in some cases provide useful genes. Usually, however, it’s maladaptive.
We observe a decrease in the frequency of archaic ancestry in the Yoruban populations in more constrained regions of the genome, suggesting that these archaic alleles have been subject to the effects of purifying selection similar to the deleterious consequences of Neanderthal and Denisovan alleles in the modern human genetic background. On the other hand, we find several loci that harbor archaic haplotypes at elevated frequencies (>60%). (Sankararaman and Durvasula 2018)
Similarly, Yang and Fu (2018) note that a "gradual decline in archaic ancestry in Europeans dating from ~37 to 14 ka suggests that purifying selection lowered the amount of Neanderthal ancestry first introduced into ancient modern humans."
This
pattern is consistent with findings from nonhuman species. A study of admixture
in trout found sharp declines in fitness even with 20% admixture. The decline
has two causes:
Hybridization can reduce fitness by either introducing alleles to a population that are not suited to the local environment (extrinsic outbreeding depression) or disrupting co-adapted gene complexes (intrinsic outbreeding depression) (Templeton 1986). These mechanisms are not mutually exclusive, and identifying the contribution of each effect is difficult. However, the high reproductive success of F1 hybrids relative to post-F1 hybrids with similar amounts of admixture suggests that some of the outbreeding depression is intrinsic. (Muhlfeld et al. 2009)
By disrupting co-adapted gene complexes, introgression causes individual genes to lose their adaptive value. Selection will thus eliminate either the introgressed alleles or the previously existing ones. In the second scenario, the complex of co-adapted genes is replaced with a simpler version.
Conclusion
Something
“clicked” in eastern Africa 80,000 to 60,000 years ago. A relatively small
group of humans acquired a new way of imagining themselves, each other, and the
world around them, and this innovation gave them an edge over everyone else.
The result: a “big bang” of population growth. They began to spread outward,
first within Africa and then into Eurasia.
Their
expansion within Africa seems to have proceeded more slowly than in Eurasia.
Initially, these modern humans were replacing “near-moderns”—people fairly
similar in appearance and genetic makeup. As they pushed farther east and
south, however, they encountered populations that were much less similar. West
Africa seems to have been home to a people who were as different from modern
humans as Neanderthals were, perhaps being related to them. In southern Africa,
modern humans encountered people even more divergent: a relic Homo erectus population. Even these
highly divergent archaic groups were not rapidly replaced; they may have
persisted as late as 15,000 years ago in West Africa and 30,000 years ago in
central Africa. Thus, modern and archaic groups seem to have long coexisted in
parts of Africa.
In
general, archaic admixture reduced fitness: “archaic alleles that introgressed
into the Yoruban population were deleterious on average”; neutral alleles were
more likely to be retained than those that had functional impacts (Sankararaman
and Durvasula 2018). A few, however, seem to have been favored by selection.
This is the case with alleles located at a tumor suppressor gene, a gene
involved with hormone regulation, and a gene involved with potassium channels.
These are individual genes, however, and it is hard to know the impact on
co-adapted gene complexes. In theory, archaic admixture should have had a
disruptive effect.
Present-day
Africans thus have admixture from a range of archaic groups, some being similar
to modern humans and others more like Neanderthals or even Homo erectus. This admixture is highest in western and southern
Africa and lowest in eastern Africa. In West Africa, admixture from a
Neanderthal-like group is estimated at 7.9% by Sankararaman and Durvasula
(2018) and at 9 to 13% by Skoglund et al. (2017). Admixture from “near-moderns”
is harder to measure. There seems to be a 13% pan-African admixture from a
population that had expanded across much of the continent some 111,000 years
ago and which perhaps spilled into the Middle East, giving rise to the
Skhul-Qafzeh hominins, i.e., early modern humans with Neanderthal culture (Watson
et al. 1997, see L1i in Table 2).
References
Bada,
J.L., R.A. Schroeder, R. Protsch, & R. Berger. (1974). Concordance of
Collagen-Based Radiocarbon and Aspartic-Acid Racemization Ages, Proceedings of the National Academy of
Sciences (USA) 71: 914-917.
http://www.pnas.org/content/71/3/914.short
Brown,
Kyle S.; Marean, Curtis W.; Jacobs, Zenobia; Schoville, Benjamin J.; Oestmo,
Simen; Fisher, Erich C.; Bernatchez, Jocelyn; Karkanas, Panagiotis; Matthews,
Thalassa (2012). An early and enduring advanced technology originating 71,000
years ago in South Africa. Nature 491 (7425):
590.
https://www.researchgate.net/publication/233331522_An_early_and_enduring_advanced_technology_originating_71000_years_ago_in_South_Africa
Dienekes
(2018). Statistical Palaeoafricans, Dienekes'
Anthropology Blog, March 25
http://dienekes.blogspot.ca/2018/03/statistical-palaeoafricans.html
Durvasula,
A., and S. Sankararaman. (2018). Recovering signals of ghost archaic admixture
in the genomes of present-day Africans, BioRxiv,
March 21
https://www.biorxiv.org/content/early/2018/03/21/285734
Hammer,
M.F., A.E. Woerner, F.L. Mendez, J.C. Watkins, and J.D. Wall. (2011). Genetic
evidence for archaic admixture in Africa, Proceedings
of the National Academy of Sciences (USA) 108: 15123-15128.
http://www.u.arizona.edu/~flmendez/papers/Hammer_2011.pdf
Harvati,
K., C. Stringer, R. Grün, M. Aubert, P. Allsworth-Jones, C.A. Folorunso.
(2011). The Later Stone Age Calvaria from Iwo Eleru, Nigeria: Morphology and
Chronology. PLoS ONE 6(9): e24024.
doi:10.1371/journal.pone.0024024
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0024024
Hawks,
J. (2012). Which population in the 1000 Genomes Project samples has the most
Neandertal similarity? John Hawks Weblog,
February 8
http://johnhawks.net/weblog/reviews/neandertals/neandertal_dna/1000-genomes-introgression-among-populations-2012.html
Hawks,
J., G. Cochran, H.C. Harpending, and B.T. Lahn. (2007). A genetic legacy from
archaic Homo, Trends in Genetics
24(1): 19-23
https://s3.amazonaws.com/academia.edu.documents/46568403/A_genetic_legacy_from_archaic_Homo20160617-13508-1nt7amy.pdf?AWSAccessKeyId=AKIAIWOWYYGZ2Y53UL3A&Expires=1522259324&Signature=Ng%2FyAWbzdtcjUJ%2BDAJTY7yE8FuA%3D&response-content-disposition=inline%3B%20filename%3DA_genetic_legacy_from_archaic_Homo.pdf
Hsieh,
P., A.W. Woerner, J.D. Wall, J. Lachance, S.A. Tishkoff, R.N. Gutenkunst, and
M.F. Hammer. (2016). Model-based analyses of whole-genome data reveal a complex
evolutionary history involving archaic introgression in Central African
Pygmies. Genome Research 26(3):
291-300
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4772012/
Irish,
J.D. (1998). Ancestral dental traits in recent Sub-Saharan Africans and the
origins of modern humans, Journal of
Human Evolution 34: 81-98.
https://www.sciencedirect.com/science/article/pii/S0047248497901913
Malekfar,
L. (2012). An analysis of the Klasies River hominins using a hybrid model, American Journal of Physical Anthropology,
Program of the 81st Annual Meeting of the American Association of Physical
Anthropologists, p. 201.
https://s3.amazonaws.com/academia.edu.documents/33649075/anthro_poster_lily_%281%29.pdf?AWSAccessKeyId=AKIAIWOWYYGZ2Y53UL3A&Expires=1522334949&Signature=h10VLn2RDPu8HtH4X55L64S2ODU%3D&response-content-disposition=inline%3B%20filename%3DPoster_An_Analysis_of_the_Klasies_River.pdf
Mayr,
E. (1970). Populations, Species, and
Evolution, Belknap Press: Cambridge (Mass.)
Muhlfeld,
C.C., S.T Kalinowski, T.E. McMahon, M.L.
Taper, S. Painter, R.F. Leary, F.W. Allendorf. (2009). Hybridization rapidly
reduces fitness of a native trout in the wild, Biology Letters, March 18
http://rsbl.royalsocietypublishing.org/content/early/2009/03/13/rsbl.2009.0033.short
Sankararaman,
S., S. Mallick, N. Patterson, and D. Reich. (2016). The combined landscape of
Denisovan and Neanderthal ancestry in present-day humans, Current Biology 26(9): 1241-1247.
https://www.sciencedirect.com/science/article/pii/S0960982216302470#bib1
Skoglund,
P., J.C. Thompson, M.E. Prendergast, A. Mittnik, K. Sirak, et al. (2017).
Reconstructing Prehistoric African Population Structure, Cell 171(1): 59-71
http://www.cell.com/cell/fulltext/S0092-8674(17)31008-5
Stojanowski,
C.M. (2014). Iwo Eleru's place among Late Pleistocene and Early Holocene
populations of North and East Africa, Journal
of Human Evolution 75: 80-89.
http://www.sciencedirect.com/science/article/pii/S0047248414000876
Stringer,
C. (2011). The chronological and evolutionary position of the Broken Hill
cranium. American Journal of Physical
Anthropology 144(supp. 52): 287
Watson,
E., P. Forster, M. Richards, and H-J. Bandelt. (1997). Mitochondrial footprints
of human expansions in Africa, American
Journal of Human Genetics 61: 691-704. 0024024
https://www.sciencedirect.com/science/article/pii/S000292970764333X
Yang,
M.A., and Q. Fu. (2018). Insights into Modern Human Prehistory Using Ancient
Genomes, Trends in Genetics 34(3):
184-196
https://www.sciencedirect.com/science/article/pii/S016895251730210X