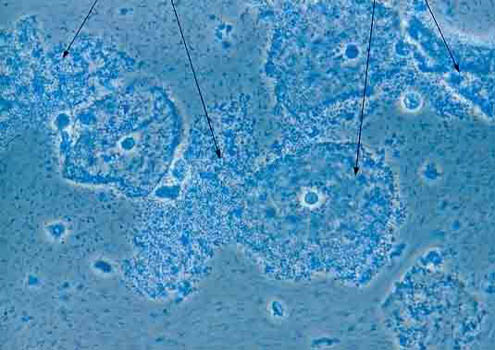
Squamous cervix cells covered with rod-shaped
bacteria, Gardnerella vaginalis
(source)
Bacterial vaginosis is a common disease among
reproductive-aged women:
[It] is characterized by the loss
of normal vaginal flora, predominantly hydrogen peroxide-producing Lactobacillus spp., and the increase in
the number and species of other bacteria in vaginal fluid. The decrease in
lactobacilli and increase in numerous facultative and anaerobic bacteria, some
of which have only been recently characterized, may lead to changes in the
characteristics of vaginal fluid, such as thin discharge and odor. (Koumans etal., 2007)
Its incidence correlates with the number of lifetime
sex partners, and this correlation holds true even when one controls for a
series of socioeconomic variables: poverty, smoking, body mass index, douching
frequency, education, and oral contraceptive use (Koumans et al., 2007). It
thus seems to be sexually transmissible, with suspicions falling particularly
on the bacterium Gardnerella vaginalis:
Sexual activity is a risk factor
for BV, and most experts believe that BV does not occur in women who have never
had vaginal intercourse [12,13].
Epidemiologic studies are strongly supportive of sexual transmission of BV
pathogens. There is a high occurrence of BV and concordance of flora in women
who have sex with women, further suggesting sexual transmission is important in
this setting [14-16]. It
is not clear, however, whether one type of sexual activity may be more
important in the pathogenesis of infection than another. As an example,
oral-genital sex may be a more important risk factor than penile intromission
into the vagina. (Sobel, 2012)
The incidence of bacterial vaginosis also correlates
with ethnicity, being 51.4% of non-Hispanic blacks, 31.9% of Mexican Americans,
and 23.2% of non-Hispanic whites. This correlation likewise holds true when the
above socioeconomic variables are held constant. High incidences have also been
found in sub-Saharan Africa (Pepin et al., 2011).
After studying these ethnic differences in vaginal
flora, Ravel et al. (2010) concluded that they were normal and not
pathological:
From these data we conclude that
vaginal bacterial communities not dominated by species of Lactobacillus are
common and appear normal in black and Hispanic women. The data from this study
are in accordance with the results of Zhou et al. (17, 18), who studied the
vaginal bacterial communities of white, black, and Japanese women. The reasons
for these differences among ethnic groups are unknown, but it is tempting to
speculate that the species composition of vaginal communities could be governed
by genetically determined differences between hosts. These might include
differences in innate and adaptive immune systems, the composition and quantity
of vaginal secretions, and ligands on epithelial cell surfaces, among others.
The ultimate cause may be vaginal pH, which is
higher in blacks and Hispanics than in Asians and non-Hispanic whites (Ravel etal., 2010). Or it may be differences in cytokine concentrations, with
differences in vaginal pH being due to the differences in vaginal flora
(Nomelini et al., 2010).
There has thus been a co-evolution between the vaginal environment and certain strains of vaginal bacteria. This co-evolution would have followed different trajectories in different human populations. In a monogamous population, possibilities for sexual transmission would have been sporadic and difficult to sustain. The picture is different in a population with a high incidence of polygyny, especially if the males often inherit or steal some of their co-wives from other males. Such a context would have favored bacteria that can spread from one co-wife to another and then to other sets of co-wives when circumstance permit.
But why wait for the right circumstances? Why not
make them by manipulating the host’s behavior? Such behavioral manipulation
sounds like science-fiction, yet it has been demonstrated in a wide range of
animal species, often in surprisingly precise ways. So how could our bacterium manipulate
its host? It wants to hop from one set of co-wives to another, but the regular
male partner is standing in its way. What should it do?
First, it should facilitate female-to-female
transmission among the co-wives. Second, it should disable the male’s
propensity for mate guarding. Better yet, it should reverse the polarity,
causing him to feel not jealousy but pleasure at the idea of being cuckolded.
This kind of manipulation occurs in the isopod Caecidotea intermedius. A parasite, Acanthocephalus dirus, infects it as a
temporary host before infecting one of several freshwater fishes. When the
parasite is still soft and immature, it cannot survive a fish eating its isopod
host. It thus seeks to reduce this risk by suppressing conspicuous host behaviors,
like mate guarding. Later, when the parasite becomes hard and mature, it can
survive consumption of its host and now stimulates mate guarding (Galipaud et
al., 2011; Mormann, 2010).
Cuckoldry is fatal to reproductive success, so any
such tendency would soon flush itself out of the gene pool. For example, the
neural networks for mate guarding might become more insensitive to outside
tampering. This change, however, would in turn favor those parasites that could
maintain such tampering. The eventual outcome would be an evolutionary
compromise where mate guarding is impaired, but not enough to prevent
reproductive success. The situation is different, though, if the parasite
spreads to another population that has never developed such immunity.
Many sexual fetishes have been around for a long
time and are often traceable to the ancient Greco-Roman world. Cuckold envy,
however, seems relatively recent, the oldest references dating back to 17th
century England (Kuchar, 2011, pp. 18-19). We may thus be looking at a sexually
transmitted parasite that entered England with the expansion of world trade in
the 17th century. But from where? Probably from a highly polygynous culture
area, like West Africa.
This parasite might be a vaginal bacterium that
first acquired sexual transmissibility and then an ability to manipulate host
behavior. It might alternately be a strain of vaginal yeast. Indeed, vaginal
strains of Candida albicans show a
similar adaptation to sexual transmission via the partner’s mouth, i.e., they adhere
better to saliva-coated surfaces than do other strains (Schmid et al., 1995).
References
Gaulipaud, M., Z. Gauthey, and L. Bollache. (2011).
Pairing success and sperm reserve of male Gammarus pulex infected by
Cyathocephalus truncatus (Cestoda: Spathebothriidea), Parasitology, 138, 11,
1429-1435.
Koumans E.H., M. Sternberg, C. Bruce, G. McQuillan, J.
Kendrick, et al. (2007). The prevalence of bacterial vaginosis in the
United States, 2001–2004; associations with symptoms, sexual behaviors, and
reproductive health, Sexually Transmitted
Diseases, 34, 864–869.
http://journals.lww.com/stdjournal/Fulltext/2007/11000/The_Prevalence_of_Bacterial_Vaginosis_in_the.6.aspx
Kuchar, G. (2001). Rhetoric, Anxiety, and the
Pleasures of Cuckoldry in the Drama of Ben Jonson and Thomas Middleton, Journal of Narrative Theory, 31 (1), Winter, pp. 1-30.
Mormann, K. (2010). Factors influencing parasite-related suppression of mating behavior in
the isopod Caecidotea intermedius, Theses and Disserations, paper 48
http://via.library.depaul.edu/etd/48
Nomelini, R.S., A.P.B. Carrijo, S.J. Adad, A.A.
Nunes, E.F.C. Murta. (2010). Relationship between infectious agents for
vulvovaginitis and skin color, Sao Paulo
Medical Journal, 128, 348-53
http://www.scielo.br/scielo.php?pid=S1516-31802010000600007&script=sci_arttext
Pépin J., S. Deslandes, G. Giroux, F. Sobéla, N.
Khonde, et al. (2011). The Complex Vaginal Flora of West African
Women with Bacterial Vaginosis. PLoS ONE,
6(9): e25082.
doi:10.1371/journal.pone.0025082
http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0025082
Ravel J, Gajer P, Abdo Z, Schneider GM, Sara S, et
al. (2010). Vaginal microbiome of reproductive-age women. Proceedings of the National Academy of Sciences U SA, 108, 4680-4687.
http://www.pnas.org/content/108/suppl.1/4680.short
Schmid, J., P.R. Hunter, G.C. White, A.K. Nand, and
R.D. Cannon. (1995). Physiological traits associated with success of Candida
albicans strains as commensal colonizers and pathogens, Journal of Clinical Microbiology, 33, 2920–2926.
Sobel, J.D. (2012). Bacterial vaginosis, Wolters
Kluwer, UpToDate
http://www.uptodate.com/contents/bacterial-vaginosis




31 comments:
So how to get rid of it?
There are sources that witness cuckoldry, or polyandry way before the 17th Century amongst the Britains.
http://www.perseus.tufts.edu/hopper/text?doc=Perseus%3Atext%3A1999.02.0001%3Abook%3D5%3Achapter%3D14
You got be kidding, right? What possible mechanism could such bacteria create to interfer with neural networks?
Theres something like 42 different bacterial strains in the vagina, so are we to presume they all control the host. Lichen Sclerosus is often associated with bacterial vaginosis and children and virgins suffer from this condition.
You got be kidding, right? What possible mechanism could such bacteria create to interfer with neural networks?
From Chapter 4, "Arms Races and Manipulation," of The Extended Phenotype, by Richard Dawkins:
"Those who have never been brainwashed or addicted to a drug find it hard to understand their fellow men who are driven by such compulsions. In the same naive way we cannot understand a host bird's being compelled to feed an absurdly oversized cuckoo, or worker ants wantonly murdering the only being in the whole world that is vital to their genetic success. But such subjective feelings are misleading, even where the relatively crude achievements of human pharmacology are concerned. With natural selection working on the problem, who would be so presumptions as to guess what feats of mind control might not be achieved? Do not expect to see animals always behaving in such a way as to maximize their own inclusive fitness. Losers in an arms race may behave in some very odd ways indeed. If they appear disoriented and unsure of their footing, this may be only the beginning."
So it seems, interestingly, to compliment Greg Cochran's infection theory of male homosexuality, it seems female bisexuality may also result from an infection. The traits of female bisexuals seems consistent with the idea that they are trying to spread some sort of bacterium between women and to men.
The plot thickens...
Wouldn't this bacteria find it easier to spread by stimulating male promiscuity instead? That would be simpler.
Cuckoldry is more easily explained as latent homosexuality
"Cuckold envy, however, seems relatively recent, the oldest references dating back to 17th century England"
Slaving in Africa goes back 3500 years and some of the groups involved were involved for the entire 3 1/2 millenia and not just the few centuries the English were.
I think the theory would be much more plausible if there were earlier examples from groups who'd been involved in black slavery for millenia e.g. Arabs, Egyptians, Phoenicians or others.
You'd imagine the groups who were involved with African slavery for the longest amount of time to have the highest incidence.
"Wouldn't this bacteria find it easier to spread by stimulating male promiscuity instead? That would be simpler."
I suppose the idea is what if a bug that evolved among a highly promiscuous r-type population with a culture that involved a lot of short-term partners came into contact with different cultures which for example
- were polygamous but with more exclusive and fewer partners
- were monogamous
- had a taboo against anal sex
then how could it adapt?
You'd imagine the groups who were involved with African slavery for the longest amount of time to have the highest incidence.
Not necessarily. Longer disease experience can lead to relative stability and greater relative immunity.
"Not necessarily. Longer disease experience can lead to relative stability and greater relative immunity."
Fair enough so theoretically it could have happened at other times and places but they became immune over time in which case it would need to have been written down before the immunity kicked in.
So you'd need a population with a written language and fairly recent exposure with records that survive.
Anon,
Polyandry is attested in many human populations, notably in Tibet and certain groups in India. It typically happens when a man doesn't have enough resources to support a wife and children, so he doubles up with one of his brothers. That way, he has some kinship with all of his children.
Cuckold envy is something else altogether. It's a feeling of sexual pleasure from visual evidence of being cuckolded. It's now a common form of sexual deviancy, yet there seems to be no evidence of it in ancient times.
Yes, there were black slaves in the Greco-Roman world, but they were only a small proportion of the total slave population. Also, most of them came from east and northeast Africa, notably Nubia, and not from West Africa. This pattern continued as the black slave trade gradually expanded during Late Antiquity and then the Islamic era. West Africa remained a secondary source of slaves for the Middle East.
Johnycomelately,
Most strains of vaginal bacteria seem to be harmless. Only a few have evolved sexual transmissibilty, but once that step is taken the possibilities for behavioral manipulation start to open up.
Jayman,
I agree. Unfortunately, most of the studies have focused on male homosexuality. And the idea of a pathogenic origin is seldom entertained.
Anon,
Cuckold envy is latent homosexuality? Don't follow you.
Anon,
See previous answer. Black slavery outside Africa did not start to become significant until Late Antiquity, and most slaves of that "first wave" came from northeast Africa, not West Africa.
Anon,
Pathogens typically develop in populations where the pool of infectable humans is quite large. For instance, South China is a place where chicken viruses can evolve into human viruses because large numbers of South Chinese farmers live in close contact with chickens.
But once that initial evolutionary change is complete, the new pathogen can spread to other populations where that initial stage is impossible. The same reasoning should apply to the evolution of STDs. They seem to evolve in highly polygynous human populations and then spread to other humans.
A Cuckold Envy bug would enhance modes of transmission rather than creating them. As female-female sex, including oral-genital sex, is likely to be less selected against in a polygynous mating system (a corollary of sexual selection of males), there is no reason to think a parasite would have to do all that much manipulation to produce lesbianism in such a population.
A CE fetish bug eliminates an obstacle to 'sexy son' genes spreading through the cuckolding of sub standard males. By reducing mate guarding by the dangerous and aggressive males, the pure 'sexy son' genes can spread without the need for genes for fighting, which interfere with attractiveness.
IMO the selective effect of a polygynous system could be enhanced by a CE bug. So the CE bug could aid 'sexy son' genes to spread by eliminating mate guarding by unsexy but dangerous husbands.
Cuckoldry is fatal to reproductive success of a cuckolded male, but it can be good for the reproductive success of the wife. The wife’s motivation for infidelity would be other man exerting greater sexual attraction through physical and mental traits such as good looks, symmetry masculinity and penis size which enhance the satifaction women get from cotius by triggering vaginal orgasm during penile-vaginal intercourse (‘the sexual behaviour consistently associated with indices of better physical and psychological health intimate relationship quality, and the one potentially reproductive behavior ’ Brody 2010).
Other sexual behavior (oral sex) is ‘variously uncorrelated or negatively associated with such indices’. From what I can gather on the net cuckold envy fetishists seem to be orientated towards performing oral-genital sex rather than the activity that women find most satisfying.
Cuckold envy fetishists are men who a woman would be better off cuckolding, especially in polygyny where a masculine man as the father of sons is the most important factor for outstanding reproductive success of a mother’s genes in the next generation. Mate guarding blocks pure selection for 'sexy son' genes
So I think that a CE bug could alter selection in polygyny to put far more emphasis on the more 'sexy son strategy genes, rather than ones selected for attractiveness to women and use of brute force to overpower and intimidate rival males out of guarding their mates. If that is true such a 'sexy son' type would be epitomised in the most polygynous areas, ie Senegal. I would like to hear any females' reactions to this face. Is it a particularly attractive type?
Cuckold fetishes can be adaptive in what I call the Twat Squeegee Theory of Sperm Competition - if the damage is already done, get aroused and get rid of the competitor's sperm and get your own in there.
"Black slavery outside Africa did not start to become significant until Late Antiquity, and most slaves of that "first wave" came from northeast Africa, not West Africa."
Yes you're right - didn't think of that. However it should still have popped up in North Africa, Spain and Portugal earlier imo. Maybe it did?
'''
If the gay pathogen idea is correct then it seems most likely to me it would have started out as a promiscuity bug which then mutated to cause bisexuality to increase transmission. So far so plausible.
Then you just need a reason why that bug or a version of that bug somehow created an exclusively gay variant - easier transmission through anal sex?
That one or both of these bugs could cause slightly different behaviour in women - promiscuous and bisexual rather than exclusively lesbian - seems reasonable also if the aim is to maximize transmission.
'''
Howevr i'm not seeing the bisexual wife fetish as cuckold envy though? As she's not getting pregnant from it and i assume - maybe wrongly - that the idea is to have sex with the wife after getting aroused by watching so...it kinda seems the opposite of cuckold envy to me?
I may be missing something here.
"Howevr i'm not seeing the bisexual wife fetish as cuckold envy though"
Although now i think of it if a man did it for real and his wife then contracted the promiscuity bug from the other woman then he could find himself cuckolded for real later on - lol - God forcing you to be good with genetics.
"If the gay pathogen idea is correct then it seems most likely to me it would have started out as a promiscuity bug which then mutated to cause bisexuality to increase transmission. So far so plausible.
"Then you just need a reason why that bug or a version of that bug somehow created an exclusively gay variant - easier transmission through anal sex?"
________________________________
I don't know how true this is although I've read it many places --that previous to the Stonewall episode that gay groups mark as the beginning of their "liberation," most gay men, at least those in the Western World, indulged in frottage or oral sex, not anal sex. Writers maintain that it was the rebelliousness of the post-Stonewall era that promoted that gay men do the "ultimate" as a sign of freedom and victory over the straight world. Today even, many gay males do not engage in anal sex, I'm told.
Further, these people also posit that in the ancient world it was frottage and/or oral sex that was practiced among the military and in pederast situations.
Thus, if true, a bug might not have been transmitted anally. Both frottage and oral sex would have led the hypothetical bug into the digestive and intestinal tracts, a common place for pathogens to reside and sometimes make side trips elsewhere.
Of course, the strategy of a bug thousands of years doesn't have to be its strategy today. A strain(s) of it could have developed that is resistant to whatever forces might have been operating against it at one time and it might have found alternate routes of survival.
Reminds me, gonorrhea has finally been declared resistant to our antibiotics. Great news, huh?
BTW, the first case of that total resistance was reported a while back by a doctor in CA treating gays.
Will they never learn?
Anon, you're getting carried away with the strategy angle, look at HIV, a known sexually transmitted bug. Exposure to porn not parasite manipulation causes HIV to spread in western hetrosexuals. A recent survey found a third of women admited to having had anal sex by age 24. That's where a lot of the 'hetrosexual' HIV infection in otherwise healthy women comes from. CCR5-Δ32, which protects against HIV, among other things, is found in 5-14% of Northern Europeans, who are not vulnerable to a bug just because it's from Africa. Europeans have not had to defend themselves against sexually transmitted diseases in a polygyous mating system, but they are formidably immunocompetent against many novel pathogens nontheless.
Having said that, it is difficult to see how cuckold envy in north Europeans could be due to cultural influences, even indirectly.
This gay man learned--and learned in time! No sex outside a monogamous relationship for me.
But yes, I expect drug-resistant gonorrhea to hit the gay male community very hard.
This gay man learned--and learned in time! No sex outside a monogamous relationship for me.
You probably mean serial monogamy. I don't think homosexuals have pure monogamous lifestyles. Serial monogamy wouldn't provide full protection. Only pure monogamy would.
That's where a lot of the 'hetrosexual' HIV infection in otherwise healthy women comes from.
I don't know if that's true. Except for blacks, heterosexuals generally have very low rates of HIV. Most of the transmission likely comes from blacks.
Anon, (too many anons BTW) Penile-vaginal intercourse is very unlikely to cause HIV infection between two otherwise healthy people, whatever their races. Some populations may be more likely to have diseases like Chlamydia which increase the chance of HIV infection.
Johnycomelately asked for a mechanism. Males performing oral genital sex would get bugs from the vagina into their mouths. Gut microbes influence behaviour .
To be even more specific, the mechanism for mate guarding behaviour involves testosterone. Gut bacteria have a profound influence on testosterone levels. Most circulating testosterone cycles through gut bacteria As Richard Jefferson points out Here. So once in the male, bugs have a quick and easy way to affect mate guarding.
Sean, thanks for the links.
From the first-
"We should start to consider the possibility that the microbiome and/or its composition may contribute to psychiatric problems."
Uh, yeah, ya think? Seems some hardheads have finally seen the light. Why the hell has it taken so many so long?
Jefferson's ideas are the opposite of Cochran's. here He says bugs evolve to help their host. Interestingly it seems the CE bug is helping the original host population to colonise a new environment.
Sean
"Anon, you're getting carried away with the strategy angle,"
That is quite possible but there is a specific reason which is the similarity in behavior between some hetero populations and homosexual men. It could be a coincidence but i wonder.
"look at HIV, a known sexually transmitted bug. Exposure to porn not parasite manipulation causes HIV to spread in western hetrosexuals. A recent survey found a third of women admited to having had anal sex by age 24."
But anal sex with who?
It's true the influence of porn has spread the fetish to white people in recent years but when i was a teenager 30 years ago the only men who ever mentioned anal sex were West Indians. It was still completely taboo otherwise.
"Black slavery outside Africa did not start to become significant until Late Antiquity, and most slaves of that "first wave" came from northeast Africa, not West Africa."
I had another thought on this.
If it was some kind of virus it might not need lots of slaves - just a single carrier - and maybe not even a slave at all but a slave trader - or not even a slave trader but a trader?
Given the ease of getting slaves from Nubia south of Egypt - (and assuming the virus if it exists is/was tropical West African) - then there would be no need to get slaves from West Africa even if the Phoenicians or Carthaginians or Sea Peoples knew of a land or sea route.
But what about gold?
http://upload.wikimedia.org/wikipedia/commons/thumb/a/a8/SONGHAI_empire_map.PNG/250px-SONGHAI_empire_map.PNG
http://en.wikipedia.org/wiki/History_of_Africa#Carthage
"The Phoenicians were seamen of the Mediterranean...trade with sub-Saharan Africa...Salt and metal goods were traded for gold, slaves, beads, and ivory."
It seems pretty far-fetched but i remember reading old Victorian history books whose explanation for the collapse of the various middle-eastern civilizations in antiquity was usually "decadence."
I always took that as Victorians being Victorians but now i wonder - was this "decadence" real and was it originally brought to those civilizations in the form of a virus?
G.Cochran cites studies showing 1% of nonpaternity, and scoffs: "I can see only one way of reconciling these equally valid kinds of data. [...] I would guess that the husband usually sets up this slightly odd situation, because of the thrill he gets in cuckolding himself. Evidently that happens frequently, not just in French farces."
Well I can think of many ways whereby it can be reconciled; women may be a lot less likely have a succesful pregnancy through sexual intercourse with a man who is not their regular partner, as Gallup suggests.
Moreover, those children (especially sons) of nonpaternity that were born may have tended not to reproduce. There is evidence from Germany for parents enhancing the mortality of their own sons, just to keep the farm together; see here. If they had strong suspicion their heir wasn't their's the father might ... correct things.
Even in those nonpaternality sons who attained adulthood, low paternal investment could have a very negative effect on their marraige prospects.
"From Chapter 4, "Arms Races and Manipulation," of The Extended Phenotype, by Richard Dawkins...."
The poster logically requested some vague mechanism demonstrating the viability of this outlandish theory. You posted a long, rambling quote from Richard Dawkins as an "answer"....even though it only offered more flighty philosophies, and nothing remotely like a mechanism.
There are far more viable explanations for the disgusting "cuckold fetish" phenomenon. (1) The boom in internet porn and the accelerating appetite for sexual novelty it generates. (2) Mass hormonal changes, such as lowered T levels, increased rates of premature ovarian failure in women leading to decreased sexual hunger in men, and other bizarre hormonal phenomena. (3) White self-hatred as fostered by pop culture and Hollywood, and pornography, could lead to many white men believing they're not "man enough" to procreate (which, indeed, they are not, if they're cuckold fetishists).
This article by Andrew Hamilton demonstrates that the anti-white messages in pornography (which tech savvy white males begin consuming at age 12 or younger) is created with racially degenerative intent. Thus, pornography is again a much more crisp and logical explanation for this: http://www.counter-currents.com/2013/08/erasing-the-line/
This is the sort of wild theory totally disconnected from mechanical practice that causes many of us to just roll our eyes at evolutionary psychology.
Accepting that sexuality is so diverse and people are driven not only by genetics, but experiences in childhood, adolescence, society...there are so many factors that contribute to our desires, arousal that have nothing to do with our identity...The concept of being a cuckold, for many men is the mental stimulation that goes along with it....yes biology can play a role in it for some men but not for all, and even among men that see themselves as a cuckold differ in views on what it is. I have published several articles on the topic and helped countless men and couples to explore their own ideas of play. Most of the time it is the man that initiates and is the person who wants to be cuckold even though, in their minds they would like it to be the woman's idea...a few women truly find themselves interested, if they can grasp the psychology behind their husbands desires to see them as having other men please them, and to dominate and humiliate their husbands, take over sexuality and tell their husband what to do....this is the ideal for the cuckold man...she gets pleasure and is satisfied but still loves him. The reality of this and her staying in a true cuckold relationship, she would have to not get emotionally attached to this other man, and given the time and her pleasure from him with repeated orgasm her biology will take over as she releases Oxytocin is a powerful hormone. When we hug or kiss a loved one, oxytocin levels drive up. It also acts as a neurotransmitter in the brain...wanting him more and her husband even less...this can become a dangerous game in a marriage...and turn into a poly relationship instead of a cuckold situation. As far as the theory of BV, that can also be transmitted from anal sex, oral sex, where there is bacteria in the mouth....a woman's own immune system, a woman having sex when she is not fully aroused and damaging her vagina, making it more susceptible to infections and bacterias. Cuckold may be a fetish or it may be an advanced erotic role play, it could be the idea of domination, or a man having problems with pleasing a woman because of a small penis or performance anxiety...it could be brought on by a women who cheated on him and through the pain he also felt a sense of arousal...the teen years are also a time when men are triggered by a sexual experiencing releasing large amounts of testosterone and it becoming a seed planted in the mind later triggered when testosterone levels drop a little and his mind now needs more mental stimulation to get that erection. I do how ever like the idea that people are exploring the topic, but as a clinical sexologist, I think that it takes on more of a justification of a problem rather than the idea of acceptance, and eroticism....
Great information. I have been suffering with BV for years and finally decided to start a blog documenting my experiences with the myriad home solutions out there. I hope it will be useful to anyone suffering from this terrible infection. Good luck :)
http://bacterialvaginosisforum.blogspot.com
No. Just no.
1) Any woman can get a yeast infection.
2) When you treat a yeast infection, you can get BV.
Or...
1) You get some other infection.
2) You get antibiotics.
3) Antibiotics throw your vaginal microflora out of whack, and you either get a yeast infection or else you can get BV directly from that.
I’m here to testify about what Dr Oseremendid for me. I have been suffering from (GENITAL HERPES VIRUS) disease for the past 4 years and had constant pain and inching, especially in my private part. During the first year, I had faith in God that i would be cured someday.This disease started circulating all over my body and I have been taking treatment from my doctor, few weeks ago I came across a testimony of Rose Smith on the internet testifying about a Man called Dr Oseremen on how he cured her from 7 years HSV 2. And she also gave the email address of this man, advise anybody to contact him for help on any kind of diseases that he would be of help, so I emailed him telling him about my (HSV 2) he told me not to worry that I was going to be cured!! Well, I never doubted him I have faith he can cure me too,, Dr Oseremen prepared and sent me Healing Oil, Soap, roots and herbs which I took. In the first one week, I started experiencing changes all over me, after four weeks of using his Roots/ Herbs, Oil and Soap, I was totally cured. no more inching , pain on me anymore as Dr Oseremen assured me. After some time I went to my doctor to do another test behold the result came out negative. So friends my advise is if you have such disease or know anyone who suffers from it or any other disease like HPV, HBV, HIV, ALS, HBP, CANCER etc. you can contact Dr oseremenfor help via dr.oseremenvadi@gamil.com or whatsapp +2349056394313
Post a Comment